
Estudo sobre alternância da Faculdade de Agricultura de Israel – Patreon
Institutos de Ciência vegetal, Faculdade de Agricultura, Universidade Hebraica de Jerusalém, Rehovot e Volcani Center, ARO, Bet-Dagan, Israel E-mail: lavee@agri.huji.ac.il
OBS.: Tradução livre e adaptação por OLIVAPEDIA

Prof Shimon Lavee
In Memoriam
Inserção imagem Olivapedia
RESUMO
O rolamento alternativo (bianualidade) é um fenômeno que ocorre com frequência em muitas espécies de árvores frutíferas e causa sérios problemas, tanto no manejo quanto na comercialização, que resultam como problemas econômicos. A oliveira mais comumente utilizada para produção de azeite e azeitona de mesa (Olea Europaea), de maneira geral, é fortemente alternada, característica definida em seu código genético. A expressão do rolamento alternativo das Oliveiras envolve uma ampla gama de mudanças na ativação e repressão do metabolismo das mesmas. (vias metabólicas endógenas). O grau de rolamento alternativo é fortemente depende das condições ambientais, e pode ser muito diferente de acordo com o clima em cada região de cultivo. O objetivo deste trabalho foi apresentar os principais fatores endógenos e ambientais e suas interações que levem a rolamentos alternativos e à revisão de abordagens com as quais o rolamento alternativo é reduzido.
Palavras-chave: Olea europaea,fisiologia, indução de broto de flor, rolamento alternativo, redução de rolamento bienal
INTRODUÇÃO
A alternância bianual ocorre em muitas espécies vegetais, o que causa dificuldades e prejuízo aos olivicultores.
Nota Olivapedia: No Brasil as espécies mais conhecidas pela afetação na alternância ad produção são as da família do café.
Os processos metabólicos e os mensageiros envolvidos ainda são pouco conhecidos, contudo não são universais e bastante diferentes entre as diversas espécies de árvores frutíferas (Goldschmidt, 2005). Embora diferentes práticas botânicas venham sendo desenvolvidas e aplicadas a fim de minimizar o rolamento bianual, os seus efeitos ainda são, na maioria dos casos, de resultado parcial (Monselise & Goldschmidt, 1982).
A oliveira domesticada (Olea europaea) é geneticamente altamente alternada na produção de azeitonas. No caso de oliveiras não irrigadas, o rendimento pode variar entre centenas de quilogramas até de 7 a 8 toneladas por hectare. A ocorrência do rolamento alternativo também ocorre fortemente em pomares com irrigação controlada, nutrição e técnicas de manejo, embora o nível de produção de frutos seja maior e melhor controlado (Lavee, 1989). Sem intervenção específica, a diferença entre anos “sem produção” e “com produção”, pode variar entre 5 e 30 T/ha… …O efeito de alternância na produção é altamente dependente das condições ambientais e pode ser muito diferente de acordo com o clima em cada região (Morettini, 1950; Hartmann, 1951). O impacto das condições ambientais não é apenas de natureza direta sobre os órgãos reprodutivos – as flores – mas também ocorre um grande impacto nos processos metabólicos endógenos da árvore. Esse impacto envolve mudanças metabólicas induzidas por ativação genética específica ou repressão.

O EFEITO DA FRUTIFICAÇÃO E DO CRESCIMENTO VEGETATIVO NA PRODUÇÃO ALTERNATIVA DE OLIVEIRAS
Para enfrentar esse problema, é necessário esclarecer primeiro o hábito frutífero das árvores. A azeitona se desenvolve a partir da inflorescência, estas por sua vez surgem apenas nos ramos brotados no ano anterior. No entanto, as inflorescências se desenvolverão apenas em madeira bem lignificada (lenhosa). Portanto, os brotos vegetativos mais fortes e mais longos crescidos no ano anterior têm um melhor potencial para desenvolver inflorescências no ano presente. Os brotos de oliveira crescem durante o verão e em muitas regiões desde muito cedo. Da primavera ao final do outono e, em casos extremos, até mesmo no inverno. Os botões que se desenvolvem nas axilas das folhas estarão, portanto, em idades diferentes na primavera seguinte, na época da floração. Ainda assim, todos os botões em partes bem lignificadas do novo ramo atingem aproximadamente o mesmo nível de desenvolvimento no final do verão e pode potencialmente se diferenciar para formar inflorescências. Em casos extremos (anos ou regiões), mesmo o crescimento muito tardio pode ser lignificado (lenhoso) e inflorescências terminais podem ocorrer.
O número de flores por inflorescência varia de 10 a 32 de acordo com o cultivar e o ano. O número potencial de flores em ano “ON” (na alternância com produção alta) extremamente alto. Um número variável de flores em cada inflorescência são flores masculinas não produtivas. O percentual das flores masculinas não afeta a produtividade, quer seja nos anos “ON”, ou nos anos “OFF”, pois apenas de 1 a 3 flores por inflorescência se tornam azeitonas, e ainda assim nem todas elas (tabela 1). Os ramos (botões) reprodutivos e vegetativos possuem a mesma origem. Uma vez que sua inflorescência se desenvolveu, um botão (ramo) reprodutivo não pode formar um rebento vegetativo, mesmo quando não deu frutos. Geralmente botões terminais vegetativos, são inibidos e desenvolvem brotos bastante fracos. Sob tais condições, a quantidade de brotos, seu comprimento e, portanto, o número de botões disponíveis para diferenciação e frutificação no ano seguinte é muito baixo. O potencial dos botões em tais brotos para se diferenciarem em botões reprodutivos é baixo, mesmo em condições ambientais favoráveis. Isso se deve a alterações metabólicas endógenas que levam à inibição da diferenciação dos botões florais em geral e flores viáveis em particular (Cuevas et al, 1994; Kitsaki et al, 1995)
O ano ‘OFF’, que resultará, é geralmente caracterizado por um estabelecimento de crescimento vegetativo vigoroso. Os brotos relativamente longos e fortes durante esse ano produzem um grande número de botões bem desenvolvidos, que sob condições ambientais adequadas estão prontos para sofrer diferenciação reprodutiva O grande número de botões que potencialmente podem se diferenciar em reprodutivos é a base para o restabelecimento de um extenso número de inflorescências, o que geralmente leva ao desenvolvimento do próximo ano ‘ON’.
Pode-se concluir que a produção de frutos na oliveira depende principalmente do crescimento vegetativo da safra anterior. Por outro lado, o grau de crescimento vegetativo em qualquer época particular é função da quantidade de frutos presentes na árvore ao mesmo tempo. Assim, o equilíbrio entre a quantidade de frutos em desenvolvimento e o crescimento vegetativo em qualquer estação de crescimento, afetará e controlará a produção potencial de frutas para a estação seguinte.

O EFEITO DO FRUTO NA INDUÇÃO DO BROTO DE FLOR
Além do efeito da fruta em geral no nível de crescimento vegetativo, os frutos em desenvolvimento mostraram-se também um efeito significativo no desenvolvimento de botões de flores para a estação seguinte. Para várias espécies de frutas foi sugerido que o fruto em desenvolvimento é forte dreno competindo nos processos metabólicos com o crescimento vegetativo (Monselise & Goldschmidt, 1982). Nas oliveiras, essa relação não é clara e parece não ser particularmente significativa.
Tabela 1: Relação entre a porcentagem de flores perfeitas e frutificação por 100 inflorescências em três cultivares de oliveira.
A, B, C = grupos de brotos com diferentes níveis de flores perfeitas dentro de cada uma das três cultivares testadas.
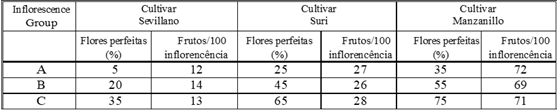
Tabela 2: O teor de proteína das folhas de oliveira maduras e da casca com um ano de idade de quatro cultivares em anos ‘ON’ e ‘OFF’ (amostras do final do verão, expresso em µg/g f.wt). Letras diferentes representam significância no nível P = 0,05.
Alguns pesquisadores encontraram uma correlação entre anos ‘ligados’ e ‘desligados’ com metabólitos primários, como carboidratos (Seyyednejad et al, 2001) e poliaminas (Pritsa & Voyiatzis, 2004). Um aumento no conteúdo de amido durante o inverno no eixo central de gemas laterais potencialmente reprodutivas também foi relatado (De la Rosa et al, 2000). Outros, entretanto, não encontraram um efeito competitivo para metabólitos orgânicos básicos entre a frutificação em desenvolvimento e o crescimento vegetativo (Fernandez-Escobar et al, 1999, 2004) Stutte & Martin (1986a) encontraram um nível uniforme de carboidratos nas folhas de árvores produtivas e não produtivas. Eles também descobriram que a destruição das sementes (embriões) em frutos intactos antes do endurecimento do caroço permitiu um desenvolvimento sem sementes dos frutos. Esses frutos “partenocárpicos” tiveram apenas um pequeno efeito no retorno da floração e no desenvolvimento da cultura no ano seguinte (Stutte & Martin, 1986b). Pode-se concluir que o efeito dos frutos em desenvolvimento na redução da diferenciação dos botões florais para a estação seguinte é de natureza regulatória por meio de sinais produzidos pelos embriões em desenvolvimento.
Tais sinais poderiam estar ligados ao desenvolvimento das diferenças significativas, encontradas no teor de proteína e suas mudanças quantitativas durante a estação de cultivo de brotos jovens de árvores frutíferas e não frutíferas (Lavee & Avidan, 1994). A mudança sazonal na quantidade de proteínas totais nas folhas e rebentos jovens encontrados durante os anos “ON” e “OFF” revelou tendências opostas (Tabela 2). Essas diferenças no conteúdo proteico parecem ser mais de natureza regulatória, pois proteínas específicas e diferentes foram induzidas durante os anos “ON” e outras nos “OFF”. A natureza dessas proteínas e seu possível papel no controle do porte alternativo são atualmente investigadas no Volcani Center em Israel.
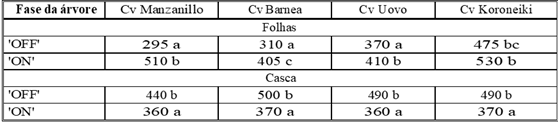
Recentemente, mudanças específicas no conteúdo mineral das folhas entre os anos “ON” e “OFF” foram relatadas e relacionadas principalmente à atividade potencial dos sistemas reguladores de crescimento (Fernandez-Escobar et al, 1999, 2004). Troncoso et al (2006) mostraram um esgotamento considerável dos teores de N e K nas folhas no final do ano “ON”, enquanto no final do ano “OFF” esses valores eram altos (Tabela 3). Eles concluíram que uma recuperação do conteúdo mineral é necessário para que a diferenciação dos botões de flores volte a ocorrer. Reguladores de crescimento e, particularmente, giberelinas1 mostraram reduzir a indução de botões de flores na oliveira, como em muitas outras espécies de frutas quando aplicadas durante a principal estação de crescimento no verão ou no outono (Lavee, 1989; Fernandez-Escobar et al, 1992; Lavee & Haskal, 1993).
No entanto, em um artigo antigo, Badr et al (1970) demonstraram que durante o inverno, pelo menos duas giberelinas, aumentaram em botões frutíferos, no entanto ácido abscísico2 foi maior no botão vegetativo. As diferenças no conteúdo de vários reguladores de crescimento nas folhas e botões durante os anos “ON” e “OFF” foram relatadas por vários pesquisadores (Navaro, 1990; Ben-Tal e Wodner, 1993; Baktir et al, 2004).
Tabela. 3: Teor de suplementos em diferentes fases do ciclo de produção.
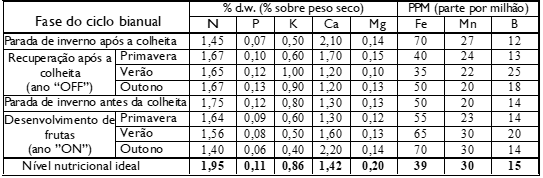
Observação Olivapedia: Para saber mais sobre a função de cada elemento químico para as plantas, acesse: Quais são os nutrientes do solo e suas funções? – OLIVAPEDIA
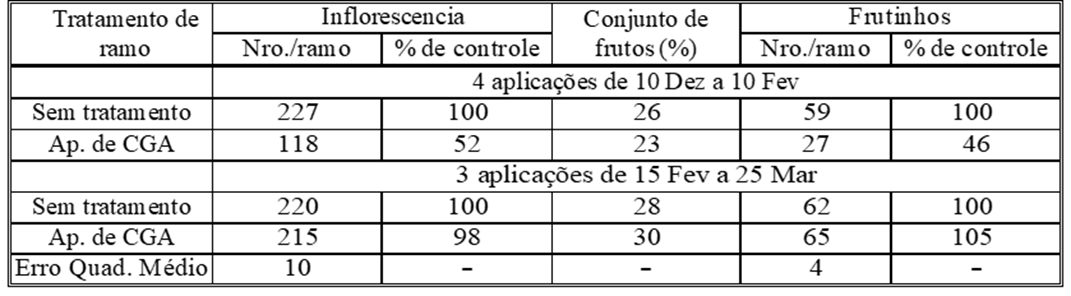
Tabela 4: O efeito das aplicações de inverno e primavera de ácido clorogênico3 exógeno (CGA) na diferenciação de botões florais e frutificação no cultivar Manzanillo. (CGA foi injetado no xilema).
As principais alterações nos vários teores de crescimento e na razão entre eles foram encontradas durante os períodos de desenvolvimento específicos. Esses períodos podem estar relacionados à diferenciação dos botões, como meados do verão, início e final de inverno e antes da abertura do botão no início da primavera (Bakir et al 2004). O envolvimento dos reguladores de crescimento e o equilíbrio entre eles regula o desenvolvimento vegetativo e frutífero atual, mas ao mesmo tempo atuam como vetor para iniciar a atividade metabólica específica controlando o potencial frutífero para o ano seguinte.
Uma mudança significativa no conteúdo de metabólitos secundários, como o ácido clorogênico nas folhas das oliveiras entre anos “ON” e “OFF”, foi notada. Supõe-se que o sinal que induz a síntese e o acúmulo desses metabólitos fenólicos nas folhas é iniciado nos embriões em desenvolvimento nos frutos. Isso poderia ser parcialmente verificado removendo as frutíferas jovens em desenvolvimento e prevenindo-as pelo aumento do nível de ácido clorogênico nas folhas, mantendo-o em um nível semelhante ao encontrado nas folhas de árvores do ano “OFF”. (Lavee & Avidan, 1981; Lavee et al, 1996; Ryan, et al, 2003) A injeção de ácido clorogênico durante o inverno antes de um ano “ON” no xilema das árvores no campo reduziu a diferenciação dos botões de flores nas estruturas tratadas em mais de 50% (Tabela 4).
Uma aplicação semelhante no início da primavera, antes da abertura do botão, não teve efeito na diferenciação do botão e no desenvolvimento das inflorescências (Lavee et al, 1986). Uma correlação positiva bastante alta foi encontrada em meados do verão entre a quantidade de frutos por árvore e o nível de ácido clorogênico em suas folhas. Conclui-se, portanto, que os frutos em desenvolvimento neste ano não estão apenas em competição com o crescimento vegetativo, mas também têm um efeito direto no metabolismo levando à indução reprodutiva e diferenciação dos botões para o rendimento potencial no ano seguinte (Fig. 1).
Geralmente, a correlação negativa entre a quantidade de produção no ano presente e a do ano seguinte já estava bem estabelecida no passado como a base para o rolamento alternativo, assim como o efeito da colheita tardia na segunda metade do inverno. No caso da colheita tardia, pode-se especular que um efeito independente adicional na inibição do crescimento do botão pode ser ativo em cima do efeito induzido anteriormente do rendimento na diferenciação do botão, levando à redução da floração e algumas vezes frutificação na primavera seguinte. A qualidade e a viabilidade das flores que se desenvolveram no ano seguinte a uma grande produção foram consideravelmente reduzidas (Cuevas et al, 1994).
EFEITO DO CLIMA NO DESENVOLVIMENTO E MANIFESTAÇÃO DO ROLAMENTOS

Em condições ambientais normais e adequadas para o desenvolvimento da oliveira, a produção alternada desenvolve-se gradualmente de forma individual em árvore. Em uma idade jovem e particularmente em boas condições de cultivo cuidados intensivos, o rendimento vai aumentando gradualmente durante os primeiros 3-4 anos do início da frutificação, e a produção alternada geralmente não se desenvolve. Posteriormente, as oliveiras começarão a alternar gradualmente em sua produção, a menos que práticas específicas de horticultura, como poda adequada, desbaste etc. sejam aplicadas. O rolamento alternativo, estando sob condições climáticas favoráveis, se manifestará de forma muito leve e muitas vezes até mesmo despercebidos para todo o pomar ou região. Abaixo de tais condições, é independente do clima, e desenvolve-se individualmente para cada árvore com base no equilíbrio entre seu crescimento individual anterior e produção de frutos.
Como resultado, a produção média do pomar poderia ser bastante uniforme com apenas pequenas flutuações por muitos anos, particularmente quando cuidados horticulturais adequados são dados às árvores. Este não será o caso em regiões onde o clima é instável e pode, em alguns anos, ser limitador, e em outros particularmente favoráveis para a diferenciação de brotos de flores e conjunto de frutas
Essas condições ocorrem principalmente nas regiões de crescimento mais quentes, onde as temperaturas de inverno (particularmente a duração e a quantidade de frio) variam significativamente de ano para ano. Regiões com geadas ocasionais da primavera, chuvas fortes ou ventos secos e quentes do deserto durante a floração também iniciarão a frutificação de forma repentina ou rolamento bianual regional. O desenvolvimento gradual do rolamento alternativo de árvores individuais é controlado por seu metabolismo endógeno, enquanto o início repentino sincronizado do pomar ou do rolamento bianual regional é induzido principalmente por fatores ambientais externos.
A temperatura é o principal fator ambiental que influencia o processo que leva à diferenciação das flores. A necessidade de quantidades adequadas de frio de inverno (abaixo de 9º) para a diferenciação dos botões das flores, foi sugerida há muitos anos (Morettini, 1950; Hartmann, 1951).
Vide também: Plantio – Parte I: Requisitos do Local – OLIVAPEDIA
O regime de temperaturas desfavoráveis, particularmente no inverno, é instrument de indução de produção alternada em geral, e em pomar ou regional sincronizada em particular. Em regiões com temperaturas de inverno uniformemente frescas, o potencial para os botões de flores se diferenciarem nas árvores é alto. Pequenas diferenças no crescimento vegetativo ou frutos a produção entre os anos levará gradualmente a um desenvolvimento alternativo de rolamentos com um cronograma individual para cada árvore. Em regiões com invernos relativamente quentes e horas marginais de frio, anos com condições de refrigeração aumentadas irão induzir uma quantidade abundante de botões reprodutivos em todas as árvores. potencialmente leva a uma produção de frutas de alta uniformidade que resultará em uma baixa diferenciação geral de botões de flores para a safra do ano seguinte e causa o início de uma orientação alternada sincronizada. Isso pode levar a uma produção de frutos alta e uniforme que resultará em uma baixa diferenciação geral dos botões de flores para a safra do ano seguinte e causará o início de uma produção alternada sincronizada. Mais comum, porém, na indução de rumo alternativo sincronizado, será a situação oposta, começando com um ano “OFF” causado, ou seja, pela falta de condições de resfriamento adequadas o suficiente (Hartmann & Prolingis, 1957).
As condições climáticas têm um grande impacto na frutificação e no desenvolvimento da oliveira em diferentes fases durante o seu ciclo de desenvolvimento anual. Nas árvores não irrigadas, a quantidade de chuva e a sua distribuição regem o nível de desenvolvimento da árvore e a seu potencial produção de frutos juntamente com a programação térmica. Qualquer condição extrema que leve a um estresse hídrico ou térmico, em qualquer estágio durante o ciclo de crescimento da árvore, pode induzir um desequilíbrio entre o desenvolvimento vegetativo e a frutificação, o que pode resultar em um início de produção alternada. Mesmo um único e curto incidente de estresse quando ocorre em um estágio crítico de desenvolvimento pode criar tal desequilíbrio e iniciar uma síndrome de rolamento bianual.
Uma iniciação mais severa de produção alternada ocorrerá em pomares irrigados e não irrigados quando a floração ou frutificação são danificadas climaticamente.
Altas temperaturas e ventos secos, por um lado, e chuvas fortes ou geadas, por outro, podem levar a uma estação infrutífera e altamente vegetativa, que por sua vez resultará em colheitas excessivas durante o ano seguinte. A produção bianual resultante pode persistir por muitos anos, se os meios de horticultura para minimizá-la não forem adotados. Em pomares irrigados, o principal parâmetro climático que afeta a produção é a temperatura, pois o estresse hídrico pode, e geralmente é evitado, pelo abastecimento de água controlado. É comumente assumido que a produção bianual em pomares intensivos é mais fraca, contudo, geralmente não é o caso. Além disso, em muitas regiões é ainda mais extremo devido ao crescimento vegetativo vigoroso nos anos “OFF” e, portanto, seguido por uma extensa produção de frutos nos “ON”, criando um ciclo vicioso de rolamento alternativo extremo.
As condições de temperatura estão intimamente envolvidas nos processos que levam à diferenciação dos botões florais e à viabilidade das flores. Em regiões de crescimento quente, invernos com resfriamento insuficiente para o desenvolvimento normal dos botões florais são bastante frequentes. A natureza do efeito da baixa temperatura no desenvolvimento dos botões reprodutivos ainda não foi resolvida. Em vários estudos, diferentes interpretações foram sugeridas para o efeito da temperatura e particularmente os baixos durante o inverno no desenvolvimento reprodutivo da oliveira. Alguns trabalhadores sugeriram que o resfriamento do inverno é necessário para liberar botões de flores predeterminados de uma fase dormente permitindo o desenvolvimento de inflorescências para uma produção futura (Rallo & Martin, 1991; Rallo et al, 1994). Outros pesquisadores associaram o efeito da exigência de frio de inverno com uma fase metabólica semelhante à vernalização4 necessária na maioria dos casos para ambos os estágios de indução e avocação da inflorescência (Hackett & Hartmann, 1967; Lavee, 1989, 1996; Troncoso et al, 2006).
A faixa de baixas temperaturas de inverno eficazes no cumprimento dos requisitos de resfriamento, sua quantidade, período indutivo e dinâmica do ciclo diário ainda não estão suficientemente claros. Em um trabalho mais antigo, Hartmann & Whisler (1975) indicaram que uma mudança gradual diária entre temperaturas baixas e altas é necessária para a diferenciação dos botões reprodutivos. Eles também mostraram que para algumas cultivares um período de temperatura uniforme moderado de 12ºC foi indutivo para a diferenciação dos botões florais. Consideravelmente mais informações são necessárias para avaliar os diferentes ciclos de temperatura diários e sazonais necessários para uma reprodução eficiente, embora balanceada, diferenciação de botões. As condições térmicas na primavera e no início do verão e a quantidade de frutos jovens em desenvolvimento na árvore estão intimamente ligados em seus efeitos sobre o nível potencial de diferenciação de botões para a produção na estação seguinte (Lavee, 1989, 1996; Fernandez-Escobar et al, 1992; Cuevas et al, 1994; Fabri & Alerci, 1999; Baktir et al, 2004). Mas, ao mesmo tempo, também é óbvio que não é o único momento para de indução, pois em muitos casos os botões que se desenvolveram consideravelmente mais tarde durante o verão e o outono também puderam ser induzidos a se diferenciar em botões de flores. Recentemente, foi demonstrado por Lavee & Troncoso (não publicado) que os botões induzidos a crescer no outono desenvolverão brotos vegetativos, enquanto os mesmos botões quando forçados posteriormente, no inverno, desenvolverão inflorescências. Como já foi indicado anteriormente, o crescimento vegetativo após invernos quentes, causando redução na diferenciação dos botões das flores, será abundante devido à falta de frutos em desenvolvimento. A produção bianual sincronizada em tais regiões é normal e comumente expressa na indústria olivícola. As temperaturas na primavera, outono e provavelmente também durante o verão estão continuamente envolvidas nos processos de indução e avocação que regem a diferenciação dos botões florais. O efeito térmico sobre a natureza do desenvolvimento do botão pode ser demonstrado em um diagrama de fluxo simples, independentemente do metabolismo endógeno envolvido (Fig. 2).
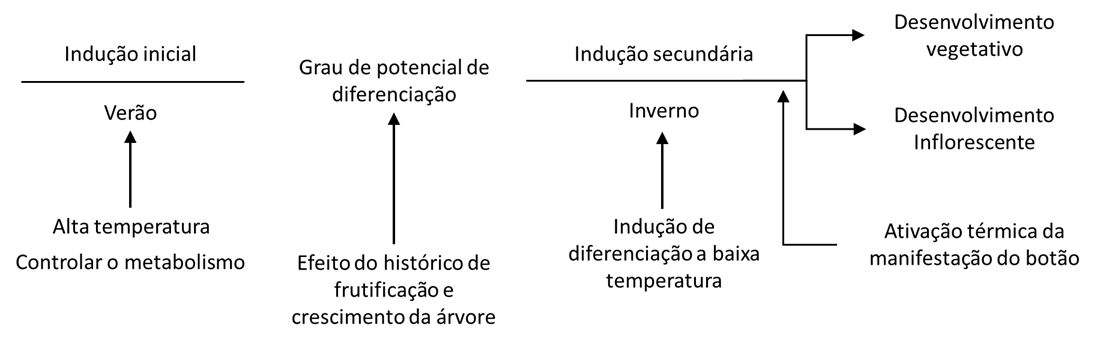
INTEGRAÇÃO DE EFEITOS ENDÓGENOS E AMBIENTAIS LEVANDO A ROLAMENTOS ALTERNATIVOS
A transformação do botão que leva aos estágios reprodutivos ou vegetativos é baseada em uma interação contínua entre as condições indutoras ambientais e a resposta metabólica endógena. Essa integração é feita resultando na expressão de diferentes níveis de crescimento e produção de frutos. O grau de expressão de fase determina as rupturas do rolamento alternativo e seu desenvolvimento a longo prazo. Como mostrado anteriormente, os fatores endógenos e ambientais estão controlando a dinâmica e o nível de cada fase de desenvolvimento da árvore, levando ao grau de desenvolvimento do rolamento alternativo. As condições ambientais e particularmente o regime de temperatura são instrumentais na ativação e repressão do gene, iniciando a atividade metabólica que leva ao desenvolvimento da fase e à expressão (manifestação) da árvore. Ainda assim, os órgãos em desenvolvimento mostraram ter um efeito de controle independente no desenvolvimento da fase futura da árvore.
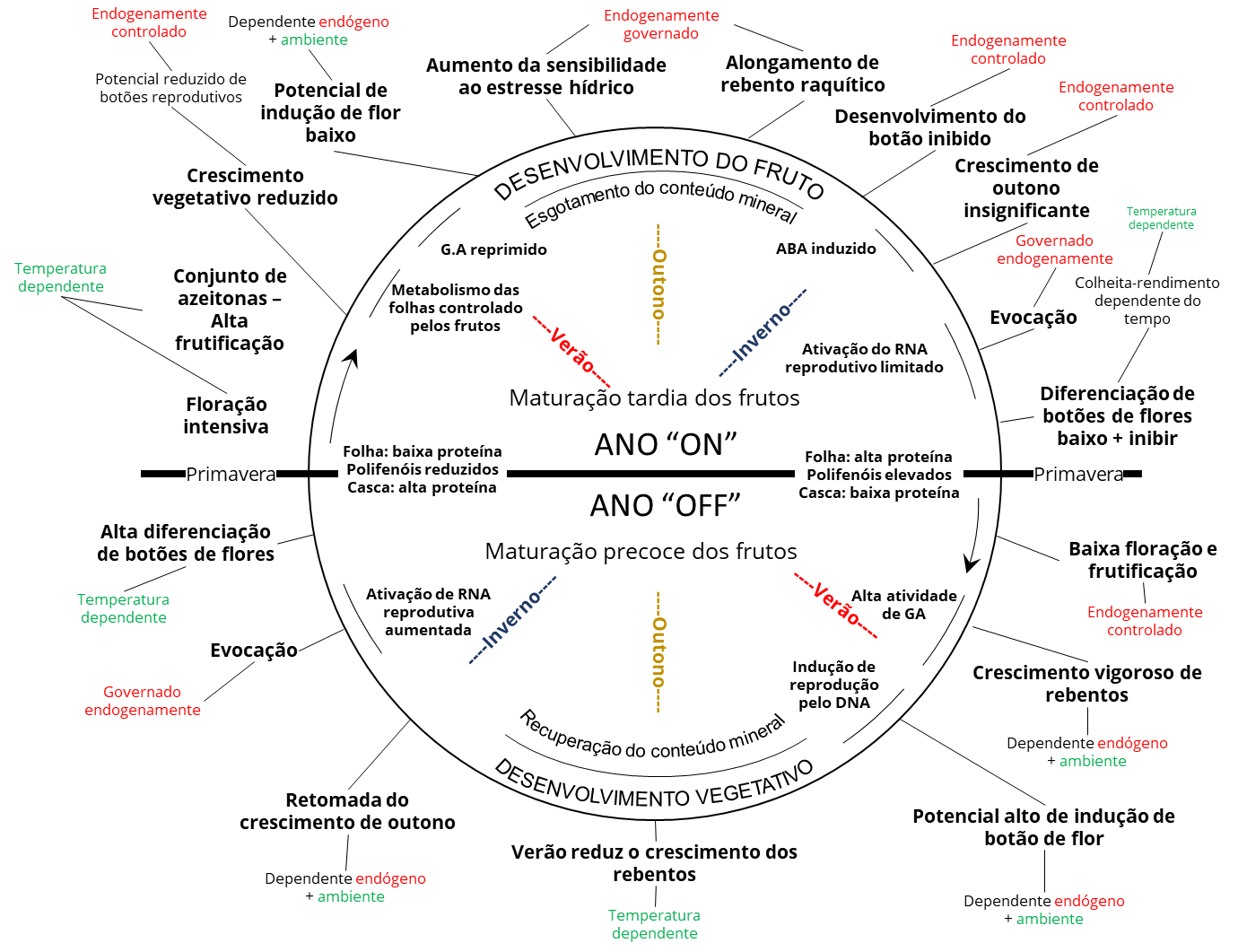
Foi feita uma tentativa de resumir o tempo e a natureza do envolvimento de fatores ambientais e endógenos durante o ciclo de desenvolvimento da oliveira em anos consecutivos “ON” e “OFF” (Fig. 3). São indicados os fatores de controle, ambiental e ou endogenamente, sobre o desenvolvimento dos órgãos. O círculo está descrevendo uma situação de rolamento alternada bianual, com a metade superior representando o ano “ligado” e a metade inferior, o “desligado”. O registro foi feito começando na primavera, a sequência de eventos durante o ano “ligado” até a primavera seguinte, de onde os eventos no ano ‘desligado’ são descritos. Ao longo do lado interno do círculo foi indicado o tempo de algumas das principais induções metabólicas. Fora do círculo, as sequências dos estágios de desenvolvimento são mostradas, bem como sua maior dependência dos efeitos ambientais, condições induzidas endógenas ou ambos.
Deve-se enfatizar que cada estágio durante o ciclo de desenvolvimento é afetado pelo anterior, mas ao mesmo tempo também afeta a expressão potencial esperada do seguinte. O nível de frutificação afetará não apenas a capacidade de diferenciação para o ano seguinte, mas também as características do fruto no presente, como tamanho do fruto, tempo de maturação do fruto e, como resultado, a taxa de acúmulo de óleo. Uma produção alta levará no ano presente uma maturação pequena e tardia dos frutos, bem como menor produção de óleo nos frutos. Nessas condições, o efeito negativo do fruto em desenvolvimento sobre a diferenciação dos botões para o ano seguinte é ampliado, enquanto a qualidade dos produtos dos frutos atuais é reduzida.
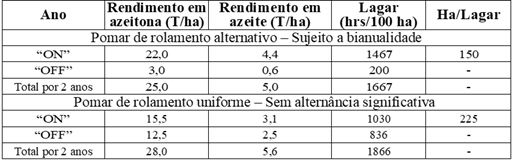
O rolamento alternativo tem, portanto, um efeito negativo não apenas na distribuição de trabalho, capacidade do moinho de óleo, necessidade de armazenamento, etc., mas também na qualidade do produto no ano “ON”. Mesmo em uma base quantitativa de dois anos, a produção de um pomar totalmente alternado está em a ordem de 10-20% menor do que uma não ou apenas parcialmente alternada (tabela 5). A questão comumente levantada de aumentar a produção alternada e ter metade do pomar produzindo em um ano e a outra metade no próximo é, geralmente, problemática. Na maioria das regiões, as condições climáticas anuais não são uniformes o suficiente para permitir um controle de produção estável a longo prazo. O rolamento se desenvolve para todo o pomar, e a maior parte das despesas economizadas em custos de cultivo e colheita (e ainda mais) tem que ser gasta em armazenamento e aumento da capacidade do lagar (moinho de óleo). Mas sobretudo, a provável redução da qualidade de pelo menos parte dos produtos da azeitona, é inviável para o mercado de hoje.
Tabela 5: Produção de frutos, rendimento de óleo e eficiência do lagar (moinho de azeite) em pomares alternados e de produção uniforme em olivais do cultivar Barnea – Os cálculos são baseados em 24 horas de trabalho e 1,5 t / h em máquinas.
ABORDAGENS E MÉTODOS PARA REDUZIR ROLAMENTOS ALTERNATIVOS
Devido à alta dependência das condições ambientais que conduzem à diferenciação dos botões florais e à frutificação, existem regiões onde a alternância da produção só pode ser parcialmente superada. Mudanças drásticas no clima, comuns particularmente na bacia do Mediterrâneo oriental, irão induzir a produção alternada bianual, mesmo após ou durante tratamentos contínuos para a regulação da frutificação. Sem uma melhor compreensão do controle molecular da diferenciação de gemas, nossa capacidade de superar a produção alternada em climas variados é geralmente temporária e incompleta. Por outro lado, em climas estáveis de forma constante, rolamentos alternativos podem ser razoavelmente bem evitados, ou continuamente minimizados. Ainda assim, um incidente inesperado pode levar a rolamentos alternativos sincronizados também em condições climáticas geralmente favoráveis. Vários métodos foram desenvolvidos e técnicas de horticultura modificadas para reduzir ou superar o rolamento alternativo.
A poda é um dos métodos básicos e mais antigos de controle da produção nos olivais. A poda tem uma ampla gama de efeitos na oliveira e nos olivais em geral. É fundamental para moldar a forma da árvore, controlar o vigor vegetativo, aumentar a penetração da luz para a regeneração e a diferenciação dos botões das flores, adaptação à pulverização e mecanização da colheita, etc.
Embora a azeitona é uma árvore setorial com cada andaime (ramificação principal partindo do tronco) atuando de forma bastante individual, algum efeito na frutificação pode ser alcançado equilibrando a quantidade de madeira para frutificação em relação à produção esperada.
A abertura das árvores para a penetração efetiva da luz na copa aumentará o potencial de frutificação, aumentando a diferenciação dos botões das flores. Ao mesmo tempo, a quantidade de “madeira” para frutificação disponível pode ser controlada. Aplicar uma poda mais severa antes do ano “ON” resultará na redução do número de frutos, limitando a quantidade de “madeira” frutífera. Também causará o início de novo crescimento vegetativo nas cunhas dos ramos podados. Este novo crescimento vegetativo pode evoluir para madeira de frutificação na estação seguinte, se uma quantidade suficiente da madeira atualmente produtiva tivesse sido removida.
Com relação ao esperado no ano “OFF”, a poda é então direcionada apenas para aumentar a penetração da luz, onde a nova copa desenvolvida é muito densa. Em regiões com um clima indutivo estável, a produção alternativa pode ser praticamente controlada pela poda. Em regiões com clima menos estável e com cultivares de crescimento lento que não respondem o suficiente à poda, métodos adicionais, como desbaste de frutos, serão necessários. As sementes em desenvolvimento neste caso, demonstraram ter um efeito negativo na diferenciação dos botões florais (Stutte & Martin, 1986b; Lavee, 1989). A redução do número de frutos nas árvores reduz o número de sementes e minimiza seu efeito inibidor na frutificação na estação seguinte. O desbaste de frutas é antes de tudo uma ferramenta para melhorar a qualidade do rendimento durante o ano “ON”, quando é realizado (Hartmann, 1952). O desbaste severo é, ao mesmo tempo, uma ferramenta útil para reduzir a produção alternativa, particularmente em regiões com produção instável. O desbaste é realizado por pulverização de ácido naftalenoacético (NAA), geralmente 10-20 dias após a plena floração, uma vez que o grau de frutificação tenha sido estabelecido (Lavee & Spiegel, 1958, 1967; Martin et al, 1980).
A aplicação de ácido giberélico (GA) no final do verão mostrou reduzir a diferenciação dos botões florais e pode ser usada para reduzir a floração nos anos “ON” (Fernandez-Escobar et al, 1992; Lavee & Haskal, 1993). A desvantagem deste método é a sua aplicação precoce, antes da indução de inverno da diferenciação dos botões florais, e das condições climáticas para a frutificação durante a floração. No norte da Itália foi relatado que uma vitamina substituta a base de NAA (“66F”) aumentou o conjunto de frutos, quando aplicado no início da floração (Bartolini et al, 1993).
Esta pode ser uma abordagem interessante para aumentar a produção de frutas no ano “OFF”. Um conjunto de frutos aprimorado também foi alcançado pela aplicação de inibidores de giberelina do grupo triazol no final do inverno, como paclobutrazol, diclorobutrazol etc., além disso causa a redução ou redução temporária do crescimento vegetativo (Prolingis & Voyiatzis, 1986; Lavee & Haskal, 1993; Rugini & Pannelli, 1993; Iannotta et al, 1999; Palliotti, 1999). O uso de Pacloburazol ou substâncias semelhantes é atualmente estudado para a regulação do crescimento e da frutificação nos novos pomares em desenvolvimento, de sebes densas e em pomares intensivos regulares antes do ano “OFF”. O tempo de colheita deve ser considerado em relação ao rolamento alternativo. Durante o estágio inicial de maturação do fruto, o tempo de colheita tem apenas um efeito muito leve na produção alternada. No entanto, a colheita tardia na maturação avançada, ou plena dos frutos, tem um efeito negativo significativo sobre as condições que levam ao desenvolvimento consecutivo da cultura uniforme. A colheita da fruta tão cedo quanto a maturação permitir, é de particular importância nos anos “ON” para evitar minimizar a redução da safra nos anos “OFF”. Evitar a colheita tardia no ano “ON” é crítico, tanto para azeite, quanto para azeitonas de mesa, mudando nos anos “ON” a colheita tardia para azeitonas colhidas antecipadamente para serem curtidas verdes.
O anelamento foi considerado um método eficiente e viável para a redução da produção alternada em olivais de mesa intensivamente cultivados. O anelamento aumentou a frutificação (Hartmann, 1905) e em algumas regiões, principalmente com invernos quentes, aumentou também o número de inflorescências quando executado antes do ano “OFF” (Lavee et al, 1983). O anelamento aumentou significativamente o número de flores perfeitas nos andaimes anelados, ampliando a proporção entre flores perfeitas e masculinas (Levin & Lavee, 2005). Para reduzir o rolamento alternativo, o anelamento de inverno é aplicado à metade dos andaimes em um ano e à segunda metade no próximo. Assim, cada andaime da árvore é anelado a cada dois anos. Este procedimento tem apenas um efeito leve e algumas vezes nenhum efeito sobre o rendimento combinado dos dois anos, mas é eficiente na redução do rolamento alternativo a longo prazo (Ben-Tal & Lavee, 1984).
Este método é particularmente útil para pomares de irrigação intensiva cultivados em regiões com condições climáticas uniformes, mesmo quando o inverno não é muito bem caracterizado. O uso contínuo do método de anelamento alternativo não tem efeito negativo no desempenho de longo prazo das árvores, desde que aplicado a plantas vigorosas, com crescimento intenso. O anelamento pode causar o declínio do andaime quando aplicado a árvores de crescimento lento e fraco, sob cultivo tradicional, sem irrigação em regiões sujeitas a estresse.
Finalmente, o possível envolvimento da nutrição, irrigação e fertirrigação no controle de rolamentos alternativos deve ser considerado. Em condições normais de crescimento equilibrado, todos os aspectos da intensificação têm muito pouca influência na produção alternada. O cultivo intensivo da oliveira aumenta a produção, mas não afeta significativamente o hábito de frutificação alternativo das árvores. Deficiências nutricionais e/ou estresse hídrico, podem aumentar a produção alternada. Nesses casos, a intervenção nutricional ou de irrigação também afetaria o nível de produção bianual. O uso desses fatores não pode ser considerado um meio significativo para reduzir rolamentos alternativos. A aplicação de fertilização e de água localmente é útil para evitar ou corrigir rolamentos alternativos em casos específico, quando foi induzido por uma deficiência nutricional aguda ou estresse hídrico, particularmente durante o período de indução inicial.
Conclusão
O rolamento alternativo é uma característica inerente às oliveiras. É principalmente controlado por uma interação entre o crescimento vegetativo e a carga de frutos. A expressão da produção alternada envolve uma ampla gama de mudanças na ativação e repressão das vias metabólicas endógenas. As condições ambientais são o principal gatilho para induzir as alterações metabólicas que levam à expressão alternada do rolamento. Uma ampla gama de eventos climáticos em diferentes estágios durante o desenvolvimento anual da oliveira pode ativar uma série de vias metabólicas relacionadas ao desenvolvimento de rolamentos alternativos.
Uma interação contínua e complexa entre a temperatura ambiente, a umidade e outros fatores ambientais estão envolvidos no desenvolvimento vegetativo e reprodutivo dos botões de oliveira. As informações sobre as vias endógenas e os genes envolvidos na diferenciação e transformação vegetativa e reprodutiva dos botões são extremamente limitadas. A natureza da transdução de sinal para iniciar o desenvolvimento vegetativo/reprodutivo balanceado ou desequilibrado da árvore ainda não é conhecida. A intervenção hortícola por meio de poda, desbaste, anelamento e outros meios culturais e nutricionais pode reduzir e até mesmo eliminar a produção alternativa em regiões com condições climáticas favoráveis e estáveis. Sob condições ambientais mais marginais e instáveis, o rolamento alternativo é mais difícil de controlar e horticultura, muitas vezes até drástica, significa que deve ser reiniciado após cada um dos vários eventos climáticos extremos. Para uma melhor compreensão e controle do rolamento alternativo, estudos em larga escala no nível molecular dos genes envolvidos na iniciação e desenvolvimento do botão da flor da oliveira e também os sinais necessários para iniciar as vias metabólicas relevantes são urgentemente necessários.
NOTAS OLIVAPEDIA:
- Giberelinas (GAs) são hormônios vegetais que regulam vários processos de desenvolvimento, incluindo alongamento de caule, germinação, dormência, floração, desenvolvimento de flores e senescência de folhas e frutos.
- Ácido abscísico é um hormônio vegetal que atua em vários processos de desenvolvimento de plantas, incluindo a dormência de sementes e gomos, o controle do tamanho do órgão e o fechamento estomático.
- Ácido clorogénico (CGA) é um ácido fenólico, éster do ácido cafeico e do ácido-quínico, por vezes designado trans-5-O-cafeoil-D-quinato, ou ácido cafeilquínico ou 5-ACQ. O composto é produzido pelas plantas, como o café e a batata, sendo que em algumas espécies é o percursor do ácido dicafeilquínico ou cianarina.
- Vernalização é uma técnica agrícola que expõe as sementes ou plantas a temperaturas baixas, o que acelera a produção de flores ou frutos. A indução floral é iniciada a partir de temperaturas entre 0ºC e 7ºC, e deve ocorrer por ciclos indutivos que podem ser de 1 a 90 dias de acordo com a espécie.
REFERÊNCIAS
Badr, S. A., H. T. Hartmann & G. C. Martin (1970): Endogenous gibberellins and inhibitors in relation to flower induction and inflorescence development in olive. Plant Physiol., 46, 674-679.
Baktir, I., S. Ulger & D. G. Himelrick (2004): Relation- ship of seasonal changes in endogenous plant hormones and alternate bearing of olive trees. HortScience, 39, 987 -990
Bartolini, S., R. Viti & C. Vitagliano (1993): Effects of different growth regulators on fruit-set in olive. Acta Hortic., 329, 246-248.
Ben-Tal, Y. S. Lavee (1984): Girdling of olive trees, a partial solution to biennial bearing II Influence of con- secutive mechanical girdling on flowering and yield. Riv. Ortoflorofrutti. Ital., 68, 441-451.
Ben-Tal, Y. & M. Wodner (1993): Involvement of different gibberellins on flowering of biennial bearing fruit trees. Bard Proj. U.S 1587/88. Report, p. 1-37.
Cuevas, J., L. Rallo & H. F. Rapaport (1994): Crop load effects on floral quality in olive. Scientia Hortic., 59, 123-130.
De la Rosa, R., L. Rallo H. F. Rapaport (2000): Olive floral bud growth and starch content during winter rest and spring budbreak. HortScience, 35, 1223-1227
Fabri, A. & L. Alerci (1999): Anatomical aspects of flower and leaf bud differentiation in Olea europaea L Acta Hortic., 474, 245-249.
Fernandez -Escobar, R., M. Benlloch, O. Navarro & G. C. Martin (1992): The time of floral induction in the olive. J. Am. Soc. Hort. Sci., 117, 304-307.
Fernandez -Escobar, R., R. Moreno & M. Garcia-Creus (1999): Seasonal changes of mineral nutrients in olive leaves during the alternate-bearing cycle Scientia Hortic,, 82, 25-45.
Fernandez -Escobar, R., R. Moreno & M. A. Sanche – Zamora (2004): Nitrogen dynamics in the olive bearing shoot. HortScience., 39, 1406-1411.
Goldschmidt, E. E. (2005): Regulatory aspects of alternate bearing in fruit trees Italus Hortus, 12, 11-17.
Hackett, W. P. & H. T. Hartmann (1967): The influence of temperature on floral initiation in the olive. Physiol. Plant, 20, 430-436.
Hartmann, H. T. (1951): Time of floral differentiation of the olive in California. Bot. Gaz., 112, 323-327.
Hartmann, H. T. (1950): The effect of girdling on flower type, fruit set and yield on the olive Proc Amer. Soc. Hort. Sci., 56, 217-226.
Hartmann, H. T. (1952): Spray thinning of olives with naphthalene-acetic acid. Proc Amer. Soc. Hort. Sci., 59, 187-195
Hartmann, H. T. & I. Prolingis (1957): Effect of different amounts of winter-chilling on fruitfulness of several olive varieties Bot. Gaz., 119, 102 104.
Hartmann, H. T. & J. E. Whisler (1975): Flower production in olive as influenced by various chilling temperature regimes. J. Amer. Soc. Hort. Sci., 100, 670-674.
Iannotta, N., L. Perri, F. Zaffina & V. Priore (1999): The influence of different types of treatment on fruit set in the Carolea cultivar (Olea europaea L.) Acta Hort , 474, 305 – 307.
Kitsaki, C. K., J. B. Drossopoulos & S. Trezis (1995): Endogenous free abscisic-acid in floral, bark and leaf tis- sues of olive during anthesis and early fruit development Scientia Hortic., 64, 95-102.
Lavee, S. (1989): Involvement of plant growth regulators and endogenous growth substances in the control of alternate bearing Acta Hortic., 239, 311-322.
Lavee, S. (1996): Biology and physiology of the olive. In: World olive Encyclopaedia. International Olive Oil Council, Madrid, p 59-106.
Lavee, S. & N. Avidan (1981): Possible involvement of chlorogenic acid in controlling alternate fruiting of the olive. Proc 13th Int Bot. Conf Sydney, p. 62.
Lavee, S. & N. Avidan (1994): Protein content and com- position of leaves and shoot bark in relation to alternate bearing of olive trees (Olea europaea L.) Acta Hortic., 356, 143-147
Lavee, S. & A. Haskal (1993): Partial fruiting regulation of olive trees (Olea europaea L ) with Paclobutrazol and Gibberellic acid in the orchard. Adv. Hort. Sci., 7, 83-86.
Lavee, S. & P. Spiegel (1958): Spray thinning of olives with growth regulators. Ktavim, 9, 129-138.
Lavee, S. & P. Spiegel-Roy (1967): The effect of time of application of two growth substances on the thinning of olive fruit. Proc Amer. Soc Hort. Sci., 91, 180-185.
Lavee, S. & M. Wodner (2004): The effect of yield, harvest time and fruit size on the oil content in fruit of irrigated olive trees (Olea europaea), cvs Barnea and Manzanillo. Scientia Hortic., 99, 267-277.
Lavee, S., H. Harshemesh & N. Avidan (1986): Phenolic acids – possible involvement in regulating growth and alternate fruiting in olive trees Acta Hortic., 179, 317-328.
Lavee, S., A. Haskal & Y. Ben-Tal (1983): Girdling olive trees, a partial solution to biennial bearing. I. Methods, timing and direct tree response. J. Amer. Soc. Hort. Sci., 58, 209-218
Levin, A. G. & S. Lavee (2005): The influence of girdling on flower type, number, inflorescence density, fruit set and yield in three different olive cultivars (Barnea, Picual and Souri). Aust. J. Agric. Res., 56, 827-831.
Martin, G. C., S. Lavee, G. S. Sibbett, C. Nishijima & S. P. Carlson (1980): A new approach to thinning olives. Calif Agric., 34(8-9), 7-8.
Monselise, P. S. E. E. Goldschmidt (1982): Alternate bearing in fruit trees Hortic Rev , 4, 1 1
Morettini, A. (1950): Olivicoltura. 1st Ed. Ramo Editoriale degli Agricoltori, Rome.
Navaro, C. (1990): Flower bud induction in Manzanillo olive. Acta Hortic., 286, 195-198.
Palliotti, A. (1999): Effect of training system on tree growth, yield and oil characteristics in different cultivars. Acta Hortic , 474, 189-192.
Prolingis, I. C. & D. G. Voyiatzis (1986): Influence of paclobutrazol plant growth regulator on vegetative and reproductive growth of olives (Olea europaea L.) Acta Hortic, 170, 587-588.
Pritsa, T. S. & D. G. Voyiatzis (2004): Seasonal changes in poly amine content of vegetative and reproductive olive organs In relation to floral initiation, anthesis, and fruit development Aust. J. Agric. Res., 1039-1046.
Rallo, L. G. C. Martin (1991): The role of chilling in releasing olive floral buds from dormancy. J Amer. Soc. Hort. Sci., 116, 1058-1062.
Rallo, L., P. Torreno, A. Vargas & J. Alvarado (1994): Dormancy and alternate bearing in olive Acta Hortic. , 356, 127-136.
Rugini, E. & G. Pannelli (1993): Preliminary results on increasing fruit set in olives (Olea europaea L.) by chemical and mechanical treatments. Acta Hortic., 329, 209- 210.
Ryan, D., P. D. Prenzler, S. Lavee, M. Antolovch & K. Robards (2003): Quantitative changes in phenolic content during physiological development of the olive (Olea europaea) cultivar Hardy’s Mammoth Int. J. Food Chem., 51, 2532-2538.
Seyyednejad, M., H. Ebrahimzadeh & A. Talaie (2001): Carbohydrate content in olive Zard c.v. and alternate bearing pattern. Int. Sugar J., 103, 84-87.
Stutte, G. & G. C. Martin (1986a): Effect of light intensity and carbohydrate reserves on flowering in olive. J Amer. Soc. Hort. Sci. , 111, 27-31.
Stutte, G. & G. C. Martin (1986b): Effect of killing seeds on return bloom of olive. Scientia Hortic. , 29, 107-113.
Troncoso, A., J. L. Garcia & S. Lavee (2006): Evaluation of the present information on the mechanisms leading to flower bud induction, evocation and differentiation 5th Int. Congress on olive growing. Turkey. Acta Hortic. (In press).